Кодон — это смысловой триплет РНК. Особенности генетического кода
Реализация генетического материала любой клетки основана на синтезе определенного набора белков, записанного в последовательности ДНК. Передача этой информации осуществляется через молекулы матричных РНК (мРНК), на основе которой строятся аминокислотные цепи. Так как белки и нуклеиновые кислоты в химическом отношении абсолютно различны, механизм комплементарного сопряжения осуществляется с участием транспортных РНК, которые взаимодействуют с матричной цепью по системе кодон-антикодон.
Особенности расшифровки последовательности мРНК
 Вам будет интересно: Созвездие Киль: характеристика и звездный состав
Вам будет интересно: Созвездие Киль: характеристика и звездный состав

Кроме разности химической природы белков и нуклеотидов в переводе генетической информации есть еще одна проблема — количественное несоответствие в разнообразии звеньев. Молекула РНК образована всего лишь четырьмя типами нуклеотидов, тогда как полипептидная цепь может включать до 20 видов аминокислот. По этой причине кодирующей единицей матрицы РНК является не один нуклеотид, а три. Эта последовательность называется триплетом.
Различные сочетания нуклеотидов в составе триплета дают 64 комбинации, что даже превышает необходимое количество вариантов, равное 20. Это явление говорит об избыточности генетического кода.
Триплетная система
Другое название смыслового триплета РНК — кодон. Эта последовательность взаимодействует с комплементарным ей антикодоном, содержащимся в молекуле транспортной РНК, которая соответствует конкретной аминокислоте. Таким образом осуществляется определяется очередность звеньев в первичной структуре белка.
Триплетная система была расшифрована в начале 1960-х годов.
Что такое кодон
Так как генетический код избыточен, некоторые аминокислоты обозначаются не одним, а несколькими кодонами. Кроме того, есть триплеты, которые вообще не содержат информацию о звене белковой последовательности. Эти кодоны нужны для остановки процесса трансляции. К ним относят UAA, UAG и UGA.
Таким образом, кодон — это состоящая из трех звеньев последовательность нуклеотидов матричной РНК, обозначающая либо аминокислоту, либо остановку трансляции. Значения всех триплетов занесены в таблицу генетического кода.
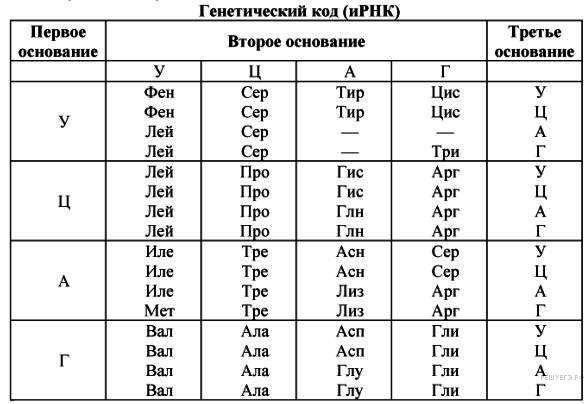
Кроме трех стоп-кодонов существует также триплет, который сигнализирует о начале транслирующего участка мРНК, — AUG. Однако, в отличие от терминирующих последовательностей, этот кодон содержит информацию об аминокислоте (метионине). Генетический код универсален для всех видов организмов.
Взаимодействие кодонов с транспортными РНК
В молекуле тРНК есть 2 функциональных участка, один из которых взаимодействует с матричной РНК, а другой связывается с аминокислотой. Антикодон содержит нуклеотиды, комплементарные последовательности кодона мРНК. Характер взаимодействия аналогичен транскрипции, только спаривание происходит группами по 3 нуклеотида.
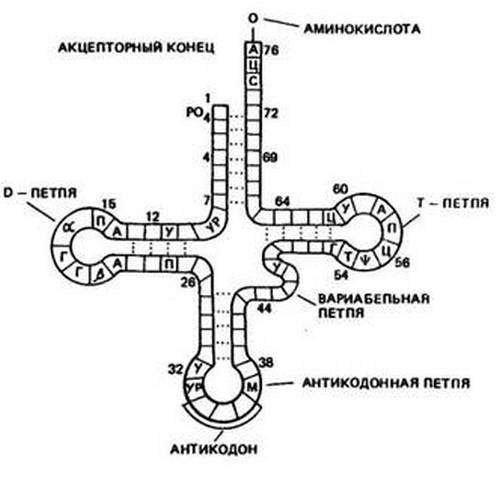
Некоторые тРНК не требуют точного комплементарного соответствия не со всеми звеньями триплета, а только с первыми двумя. Толерантность к третьему нуклеотиду в кодоне называется качанием, благодаря которому одна тРНК может связываться с несколькими видами триплетов, отличающихся друг от друга только звеном в последней позиции.
Anticodon описание, функции и различия с кодоном
антикодонной представляет собой последовательность из трех нуклеотидов, которая присутствует в молекуле переносящей РНК (тРНК), функция которой заключается в распознавании другой последовательности из трех нуклеотидов, которая присутствует в молекуле мессенджер-РНК (мРНК).
Таким образом, во время трансляции молекулы РНК-мессенджера «читаются» посредством распознавания их кодонов антикодонами переносящих РНК. Эти молекулы называются так, потому что они передают определенную аминокислоту белковой молекуле, которая образуется в рибосоме..
Есть 20 аминокислот, каждая из которых кодируется определенным триплетом. Однако некоторые аминокислоты кодируются более чем одним триплетом.
Кроме того, некоторые кодоны распознаются антикодонами в переносимых молекулах РНК, к которым не присоединены аминокислоты; это так называемые стоп-кодоны.
описание
Антикодон образован последовательностью из трех нуклеотидов, которая может содержать любое из следующих азотистых оснований: аденин (A), гуанин (G), урацил (U) или цитозин (C) в комбинации из трех нуклеотидов таким образом, что это работает как код.
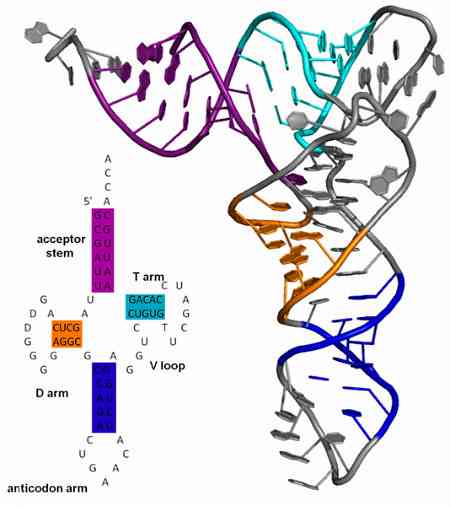
Антикодоны всегда находятся в переносящих молекулах РНК и всегда лежат в направлении 3 ‘-> 5’. Структура этих тРНК сходна с клевером таким образом, что она подразделяется на четыре петли (или петли); в одной из петель находится антикодон.
Антикодоны необходимы для распознавания кодонов РНК-мессенджера и, следовательно, для процесса синтеза белка во всех живых клетках..
функции
Поскольку транскрипция (синтез копий РНК-мессенджера) происходит в направлении 5 ‘-> 3’, кодоны в РНК-мессенджере имеют эту ориентацию. Следовательно, антикодоны, присутствующие в молекулах переносящей РНК, должны иметь противоположную ориентацию, 3 ‘-> 5’.
Этот союз обусловлен взаимодополняемостью. Например, если один кодон является 5′-AGG-3 ‘, то антикодон является 3′-UCC-5’. Этот тип специфического взаимодействия между кодонами и антикодонами является важным этапом, который позволяет нуклеотидной последовательности в РНК-мессенджере кодировать последовательность аминокислот внутри белка..
Различия между антикодоном и кодоном
— Антикодоны являются тринуклеотидными единицами в тРНК, комплементарными кодонам в мРНК. Они позволяют тРНК доставлять правильные аминокислоты во время производства белка. Напротив, кодоны являются единицами тринуклеотидов в ДНК или мРНК, которые кодируют определенную аминокислоту в синтезе белка..
— Антикодоны являются связующим звеном между нуклеотидной последовательностью мРНК и аминокислотной последовательностью белка. И наоборот, кодоны передают генетическую информацию из ядра, где находится ДНК, в рибосомы, где происходит синтез белка..
— Антикодон находится в плече Anticodon молекулы тРНК, в отличие от кодонов, которые находятся в молекуле ДНК и мРНК..
— Антикодон дополняет соответствующий кодон. Напротив, кодон в мРНК комплементарен триплету нуклеотидов определенного гена в ДНК.
— ТРНК содержит антикодон. Напротив, мРНК содержит ряд кодонов.
Гипотеза качения
Балансирующая гипотеза предполагает, что соединения между третьим нуклеотидом кодона мессенджерной РНК и первым нуклеотидом антикодона переносящей РНК менее специфичны, чем соединения между двумя другими нуклеотидами триплета..
Крик описал это явление как «раскачивание» в третьей позиции каждого кодона. В этой позиции происходит нечто, что позволяет профсоюзам быть менее строгими, чем обычно. Это также известно как колебание или тамболео.
Эта гипотеза колебания Крика объясняет, как антикодон данной тРНК может быть спарен с двумя или тремя разными кодонами мРНК..
Крик предположил, что, поскольку спаривание оснований (между основанием 59 антикодона в тРНК и основанием 39 кодона в мРНК) является менее строгим, чем обычно, в этом сайте допускается определенное «колебание» или пониженное сродство.
В результате одна тРНК часто распознает два или три родственных кодона, которые определяют данную аминокислоту.
Обычно водородные связи между основаниями антикодонов тРНК и кодонов мРНК следуют строгим правилам спаривания оснований только для первых двух оснований кодона. Однако этот эффект встречается не во всех третьих позициях всех кодонов мРНК..
РНК и аминокислоты
Основываясь на гипотезе вобуляции, было предсказано существование по крайней мере двух РНК-переносчиков для каждой аминокислоты с кодонами, проявляющими полную дегенерацию, что подтвердилось..
Эта гипотеза также предсказала появление трех переносящих РНК для всех шести сериновых кодонов. Действительно, три тРНК для серина были охарактеризованы:
— ТРНК для серина 1 (антикодон AGG) связывается с кодонами UCU и UCC.
— ТРНК для серина 2 (антикодон AGU) связывается с кодонами UCA и UCG.
— ТРНК для серина 3 (антикодон UCG) связывается с кодонами AGU и AGC.
Эти особенности были подтверждены стимулированным связыванием очищенных триауклеотидов аминоацил-тРНК с рибосомами in vitro..
Наконец, несколько переносящих РНК содержат основание инозина, которое сделано из гипоксантинового пурина. Инозин получают пост-транскрипционной модификацией аденозина.
Гипотеза Крика предсказывает, что, когда инозин присутствует на 5′-конце антикодона (позиция колебаний), он будет соединяться с урацилом, цитозином или аденином в кодоне..
Фактически, очищенная аланил-тРНК, содержащая инозин (I) в 5 ‘положении антикодона, связывается с рибосомами, активированными тринуклеотидами GCU, GCC или GCA.
Тот же результат был получен с другими тРНК, очищенными инозином в 5′-положении антикодона. Таким образом, гипотеза Крика о колебаниях очень хорошо объясняет отношения между тРНК и кодонами с учетом генетического кода, который является вырожденным, но упорядоченным..
Антикодон: описание, функции и отличие от кодона
Содержание:
А антикодон представляет собой последовательность из трех нуклеотидов, которая присутствует в молекуле РНК-переносчика (тРНК), функция которой заключается в распознавании другой последовательности из трех нуклеотидов, которая присутствует в молекуле матричной РНК (мРНК).
Это распознавание между кодонами и антикодонами антипараллельно; то есть один расположен в направлении 5 ‘-> 3’, а другой соединен в направлении 3 ‘-> 5’. Это распознавание между последовательностями из трех нуклеотидов (триплетов) необходимо для процесса трансляции; то есть в синтезе белков в рибосоме.
Таким образом, во время трансляции молекулы информационной РНК «считываются» благодаря распознаванию их кодонов антикодонами транспортных РНК. Эти молекулы названы так потому, что они передают определенную аминокислоту молекуле белка, которая образуется на рибосоме.
Есть 20 аминокислот, каждая из которых кодируется определенным триплетом. Однако некоторые аминокислоты кодируются более чем одним триплетом.
Кроме того, некоторые кодоны распознаются антикодонами в молекулах транспортной РНК, к которым не присоединены никакие аминокислоты; это так называемые стоп-кодоны.
Описание
Антикодон состоит из последовательности из трех нуклеотидов, которые могут содержать любое из следующих азотистых оснований: аденин (A), гуанин (G), урацил (U) или цитозин (C) в комбинации из трех нуклеотидов таким образом, что он работает как код.
Антикодоны всегда находятся в молекулах транспортной РНК и всегда расположены в направлении 3 ‘-> 5’. Структура этих тРНК подобна структуре клевера, так что она подразделяется на четыре петли (или петли); в одной из петель находится антикодон.
Антикодоны необходимы для распознавания кодонов информационной РНК и, следовательно, для процесса синтеза белка во всех живых клетках.
Характеристики
Поскольку транскрипция (синтез копий информационной РНК) происходит в направлении 5 ‘-> 3’, кодоны информационной РНК имеют эту ориентацию. Следовательно, антикодоны, присутствующие в молекулах транспортной РНК, должны иметь противоположную ориентацию, 3 ‘-> 5’.
Этот союз обусловлен дополнительностью. Например, если кодоном является 5′-AGG-3 ‘, антикодоном является 3′-UCC-5’. Этот тип специфического взаимодействия между кодонами и антикодонами является важным шагом, который позволяет нуклеотидной последовательности в информационной РНК кодировать аминокислотную последовательность в белке.
Различия между антикодоном и кодоном
— Антикодон находится в антикодоновом плече молекулы тРНК, в отличие от кодонов, которые расположены в молекуле ДНК и мРНК.
— Антикодон является дополнительным к соответствующему кодону. Вместо этого кодон в мРНК комплементарен триплету нуклеотидов определенного гена в ДНК.
— тРНК содержит антикодон. Напротив, мРНК содержит ряд кодонов.
Гипотеза качелей
Гипотеза качания предполагает, что соединения между третьим нуклеотидом кодона информационной РНК и первым нуклеотидом антикодона транспортной РНК менее специфичны, чем соединения между двумя другими нуклеотидами триплета.
Крик описал это явление как «раскачивание» в третьей позиции каждого кодона. Что-то происходит в этом положении, что позволяет суставам быть менее строгими, чем обычно. Это также известно как колебание или колебание.
Эта гипотеза криковского колебания объясняет, как антикодон данной тРНК может сочетаться с двумя или тремя различными кодонами мРНК.
Крик предположил, что, поскольку спаривание оснований (между основанием 59 антикодона в тРНК и основанием 39 кодона в мРНК) менее жесткое, чем обычно, на этом участке допускается некоторое «колебание» или пониженное сродство.
В результате одна тРНК часто распознает два или три связанных кодона, которые определяют данную аминокислоту.
Обычно водородные связи между основаниями антикодонов тРНК и кодонами мРНК подчиняются строгим правилам спаривания оснований только для первых двух оснований кодона. Однако этот эффект проявляется не во всех третьих положениях всех кодонов мРНК.
РНК и аминокислоты
Основываясь на гипотезе колебания, было предсказано существование по крайней мере двух РНК переноса для каждой аминокислоты с кодонами, демонстрирующими полную вырожденность, что оказалось правдой.
Эта гипотеза также предсказывала появление трех РНК переноса для шести кодонов серина. Действительно, для серина были охарактеризованы три тРНК:
— тРНК серина 1 (антикодон AGG) связывается с кодонами UCU и UCC.
— тРНК серина 2 (антикодон AGU) связывается с кодонами UCA и UCG.
— тРНК серина 3 (антикодон UCG) связывается с кодонами AGU и AGC.
Эти специфичности были подтверждены путем стимулирования связывания очищенных тринуклеотидов аминоацил-тРНК с рибосомами in vitro.
Наконец, несколько транспортных РНК содержат основание инозин, которое состоит из пуринового гипоксантина. Инозин продуцируется посттранскрипционной модификацией аденозина.
Гипотеза Крика предсказывает, что когда инозин присутствует на 5′-конце антикодона (положение колебания), он будет спариваться с урацилом, цитозином или аденином на кодоне.
Фактически, очищенная аланил-тРНК, содержащая инозин (I) в 5′-положении антикодона, связывается с рибосомами, активированными тринуклеотидами GCU, GCC или GCA.
Тот же результат был получен с другими тРНК, очищенными инозином в 5′-положении антикодона. Таким образом, гипотеза крикового колебания очень хорошо объясняет отношения между тРНК и кодонами с учетом генетического кода, который является вырожденным, но упорядоченным.
Генетический код. Биосинтез белка
теория по биологии 🌿 основы генетики
Генетическая информация и генетический код
Каждый вид имеет свой собственный, отличный от других видов, набор белков. Интересно то, что белки, выполняющие идентичные функции у разных видов могут быть похожими или даже абсолютно идентичными.
У белков есть несколько состояний их структур:
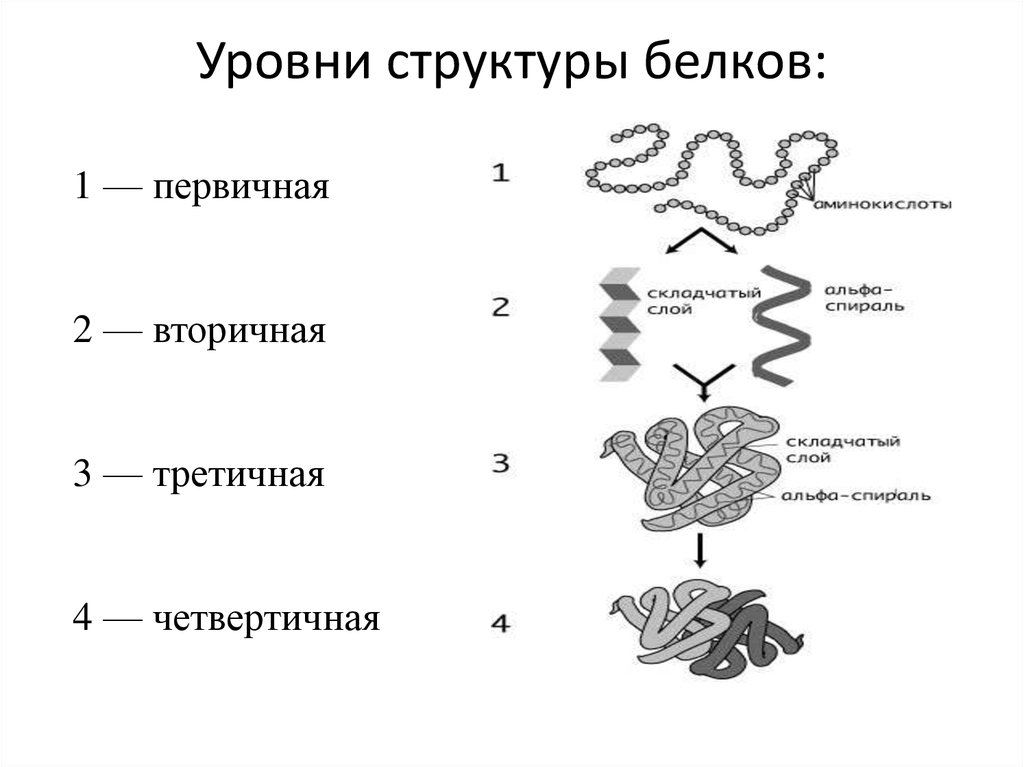
Именно первичная структура является определяющей свойства белка. Эта структура – цепь из аминокислот. Аминокислоты, в свою очередь, представляют собой ряд триплетов из нуклеотидов. Решая генетические задачи, обращаются как раз-таки к знакомой таблице:
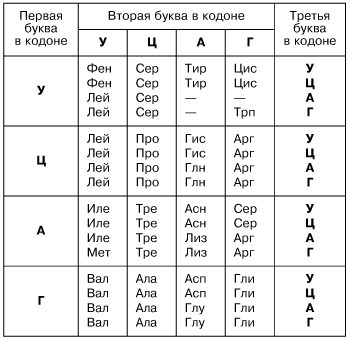
Каждая аминокислота кодируется тремя нуклеотидами, которые составляют триплет или иначе кодон. Именно последовательность нуклеотидов называется генетической информацией, а участок последовательности, в котором хранится информация о первичной структуре белка это и есть ген.
Нуклеотиды, составляющие ДНК и РНК различаются:
В состав ДНК входят:
В состав РНК входят:
Кроме того, в составе РНК (рибонуклеиновой кислоты) сахар рибоза, а ДНК (дезоксирибонуклеиновой кислоты) — дезоксирибоза. РНК — одноцепочечная, а ДНК — двухцепочечная.
Между нуклеотидами есть водородные связи. Они могут быть как двойные, так и тройные. Нуклеотиды не могу быть связаны в случайном порядке. Для этого существует принцип комплементарности ДНК, по которому аденин одной цепи ДНК соединяется с тимином другой цепи ДНК, другая пара в ДНК – гуанин – цитозин. В РНК все аналогично, за исключением того, что вместо тимина там урацил. Между парами А-Т/А-У – две водородных связи, а между парами Ц-Г – три. На письме это обозначается чёрточками: двойная связь как знак «равно», а тройная – три горизонтальные черты.
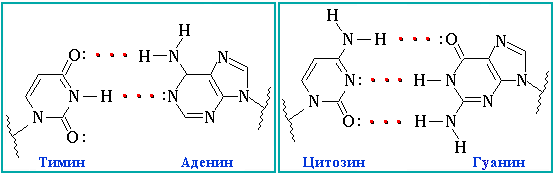
Свойства генетического кода
Транскрипция и трансляция
Из цитологии известно, что генетическая информация у эукариотических клеток заключена в ядре в виде ДНК. Однако процесс биосинтеза белка происходит в цитоплазме на рибосомах.
Спиральная цепь ДНК при раскручивается, в это время по одной из цепочек ДНК строится комплементарная цепь. Из ядра в цитоплазму информация выходит в виде информационной РНК (иРНК). иРНК комплементарная одной из цепей ДНК. Этот процесс переписывания называется транскрипцией. Полученная цепь практически идентичная другой цепи ДНК, за исключением того, что вместо тимина там урацил.В процессе участвует специальный фермент РНК-полимераза.
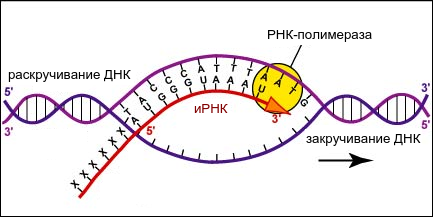
Теперь в ядре есть цепочка, которая уже начала процесс биосинтеза. Как говорилось выше, процесс ассимиляции идет на рибосомах. иРНК выходит в цитоплазму через поры ядерной мембраны
тРНК по форме напоминает лист клевера, а по принципу работы – штамп. На него, прямо как чернила, наслаиваются кодоны.
В цитоплазме начинается процесс трансляции, то есть перевод последовательности нуклеотидов информационной РНК в последовательность аминокислот белка.
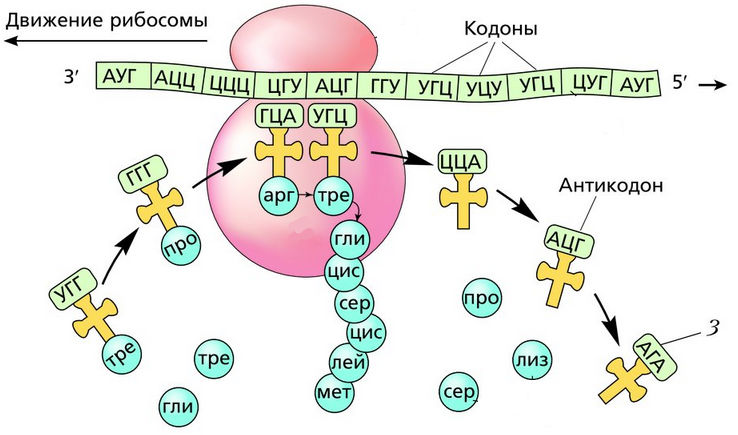
Рибосома захватывает стартовый конец цепи иРНК. Затем она начинает двигаться по цепи, одна остановка рибосомы происходит на 6-ти нуклеотидах. В это время молекула тРНК, на которых есть триплет аминокислоты «подлетает» к цепи, в месте, где находится рибосома. За время остановки рибосомы транспортная РНК успевает распознать свою пару на цепи иРНК, которая называется антикодоном. Тогда тРНК «ставит свой штамп», оставляя на цепи свой кодон. Между нуклеотидами образуются водородные связи. Так нарастает новая цепь. На одной информационной РНК работает сразу много рибосом, поэтому работа идет очень быстро. Совокупность рибосом, синтезирующих на одной иРНК, называется полисомой.
По окончанию процесса биосинтеза, цепочка отсоединяется от рибосомы и принимает свою природную структуру: вторичную, третичную или четвертичную.
pазбирался: Надежда | обсудить разбор | оценить
В современной генетической инженерии часто применняют технологии, связанные с гомологичной рекомбинацией ДНК непосредственно в живом объекте. Один из примеров – система CRE-Lox P. Lox P – это последовательность нуклеотидов в ДНК фага Р1. Она состоит из 34 нуклеотидов. В середине располагается несимметричная последовательность из 8 нуклеотидов (показана серой стрелкой на рисунке). По краям располагаются так называемые палиндромные последовательности из 13 нуклеотидов (выделены на рисунке как пунктирные блоки). Они симметричны (чтобы в этом убедиться, достаточно прочитать обе последовательность от 5´- конца к 3´- концу). Именно эти палиндромные участки узнаёт особый фермент, вызывающий рекомбинацию, который обозначают CRE. Будем в дальнейшем называть этот фермент рекомбиназой CRE. Для того, чтобы состоялась рекомбинация, два сайта Lox P должны расположиться параллельно друг другу. Рекомбиназа CRE узнает эти сайты, внесет в ДНК разрезы в определённых местах, а затем 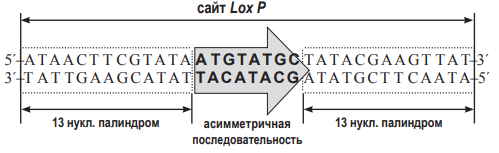 соединит по-новому две нити ДНК (т.е. произойдет рекомбинация). Аналогично работает и другая система гомологичной рекомбинации – Flp-FRT, обнаруженная у пекарских дрожжей. Сайт FRT – это последовательность ДНК, которую узнает свой фермент гомологичной рекомбинации – флиппаза (Flp).
соединит по-новому две нити ДНК (т.е. произойдет рекомбинация). Аналогично работает и другая система гомологичной рекомбинации – Flp-FRT, обнаруженная у пекарских дрожжей. Сайт FRT – это последовательность ДНК, которую узнает свой фермент гомологичной рекомбинации – флиппаза (Flp). 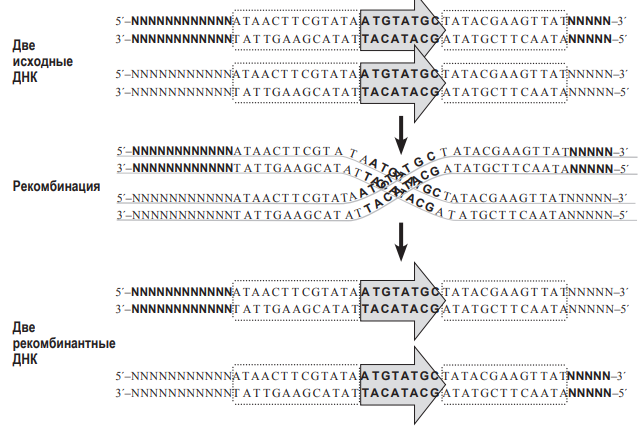 При рекомбинации две молекулы ДНК должны ориентироваться параллельно друг другу сайтами FRT, и только в этом случае произойдёт рекомбинация. Заметим, что флиппаза Flp узнает только свою последовательность FRT, но не может работать с сайтами Lox P, а рекомбиназа CRE узнает только свои сайты Lox P, но не работает с сайтами FRT. Предварительное доказательство (лемма) к задаче 9 (5 баллов). 1. Докажем, что при гомологичной рекомбинаци по «перевёрнутым» (инвертированным) повторам происходит «переворот» последовательности ДНК, находящейся между повторами. Для этого нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек.
При рекомбинации две молекулы ДНК должны ориентироваться параллельно друг другу сайтами FRT, и только в этом случае произойдёт рекомбинация. Заметим, что флиппаза Flp узнает только свою последовательность FRT, но не может работать с сайтами Lox P, а рекомбиназа CRE узнает только свои сайты Lox P, но не работает с сайтами FRT. Предварительное доказательство (лемма) к задаче 9 (5 баллов). 1. Докажем, что при гомологичной рекомбинаци по «перевёрнутым» (инвертированным) повторам происходит «переворот» последовательности ДНК, находящейся между повторами. Для этого нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек.  Затем «изогнём» молекулу так, чтобы повторы, обозначенные стрелками, встали параллельно друг другу. После обмена участками и «распрамления» окажется, что центральная часть между повторами «перевернулась».
Затем «изогнём» молекулу так, чтобы повторы, обозначенные стрелками, встали параллельно друг другу. После обмена участками и «распрамления» окажется, что центральная часть между повторами «перевернулась». 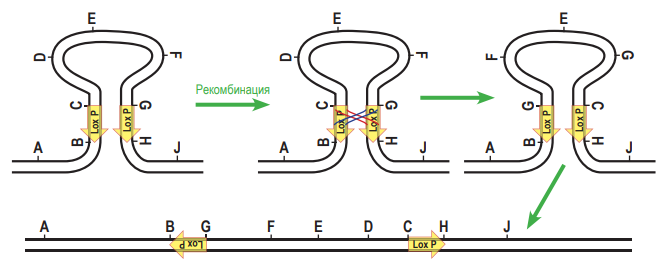 2. Докажем, что при гомологичной рекомбинаци по прямым повторам происходит образование кольцевой ДНК, при этом из линейной последовательности ДНК «удаляется» участок, находящейся между повторами. Для этого используем тот же приём: нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек. Только в этом случае для того, чтобы прямые повторы встали параллельно друг другу, придётся хитроумно изогнуть молекулу так, чтобы от конца одного из повторов (точка С) шли точки D, E, F, а потом начинался новый повтор (в точке G). После рекомбинации точки С и G поменяются местами, и в результате получится кольцевая ДНК (C, D, E, F, G) и линейный участок (A, B, H, J). Будем считать, что кольцевая ДНК как бы «исчезает» (не может реплицироваться в клетке).
2. Докажем, что при гомологичной рекомбинаци по прямым повторам происходит образование кольцевой ДНК, при этом из линейной последовательности ДНК «удаляется» участок, находящейся между повторами. Для этого используем тот же приём: нарисуем молекулу ДНК и условно обозначим на ней буквами несколько точек. Только в этом случае для того, чтобы прямые повторы встали параллельно друг другу, придётся хитроумно изогнуть молекулу так, чтобы от конца одного из повторов (точка С) шли точки D, E, F, а потом начинался новый повтор (в точке G). После рекомбинации точки С и G поменяются местами, и в результате получится кольцевая ДНК (C, D, E, F, G) и линейный участок (A, B, H, J). Будем считать, что кольцевая ДНК как бы «исчезает» (не может реплицироваться в клетке). 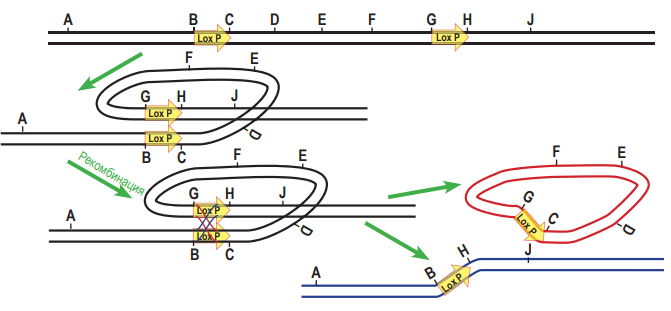 А. Поскольку после 35S-промотора на той же цепи ДНК располагается кодирующая часть гена DsRed, клетки должна светиться красным светом. Б. Рекомбиаза CRE узнаёт последовательнсоти LoxP. Если повторы расположены инвертированно, то произойдёт «переворот» последовательности ДНК, расположенной между повторами. Таким образом, после рекомбинации конструкция будет выглядеть следущим образом:
А. Поскольку после 35S-промотора на той же цепи ДНК располагается кодирующая часть гена DsRed, клетки должна светиться красным светом. Б. Рекомбиаза CRE узнаёт последовательнсоти LoxP. Если повторы расположены инвертированно, то произойдёт «переворот» последовательности ДНК, расположенной между повторами. Таким образом, после рекомбинации конструкция будет выглядеть следущим образом: 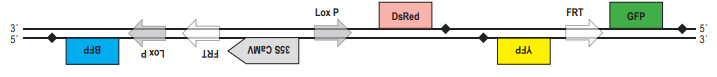 Свечение клеток изменится, поскольку после промотора на той же цепи ДНК окажется гена BFP, обестпечивающий синее свечение клеток. В. При рекомбинации по прямым повторам происходит потеря участка ДНК, расположенного между ними. Из двух повторов остаётся только один. Таким образом, после рекомбинации по сайтам FRT конструкция будет выглядеть следующим образом:
Свечение клеток изменится, поскольку после промотора на той же цепи ДНК окажется гена BFP, обестпечивающий синее свечение клеток. В. При рекомбинации по прямым повторам происходит потеря участка ДНК, расположенного между ними. Из двух повторов остаётся только один. Таким образом, после рекомбинации по сайтам FRT конструкция будет выглядеть следующим образом: 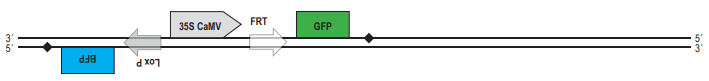 Клетки будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP. Г. После действия рекомбиназы CRE те последовательности, на которые может действовать флип паза Flp, «перевернулись», и вместо прямых стали инвертрованными. После рекомбинации участок между ними также должен «перевернуться»:
Клетки будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP. Г. После действия рекомбиназы CRE те последовательности, на которые может действовать флип паза Flp, «перевернулись», и вместо прямых стали инвертрованными. После рекомбинации участок между ними также должен «перевернуться»: 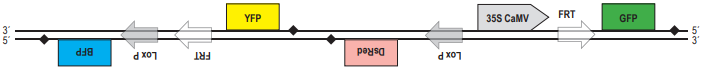 В этом случае клетки также будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP.
В этом случае клетки также будут светиться зелёным светом за счёт того, что под промотором оказалась кодирующая последовательность гена GFP.
pазбирался: Надежда | обсудить разбор | оценить
pазбирался: Надежда | обсудить разбор | оценить
Сначала найдём место расщепления плазмиды рестриктазой BglII:  Таких участков оказывается два. В результате расщепления из плазмиды выщепляется короткий фрагмент:
Таких участков оказывается два. В результате расщепления из плазмиды выщепляется короткий фрагмент:  Остаётся укороченная линейная ДНК, содержащая интактный ген устойчивости к ампицилину и расщеплённый ген устойчивости к эритромицину.
Остаётся укороченная линейная ДНК, содержащая интактный ген устойчивости к ампицилину и расщеплённый ген устойчивости к эритромицину.  При сшивании липких концов ДНК-лигазой наиболее часто будут соединяться концы этой молекулы и образовываться кольцо длиной 4163 нуклеотида. Такая ДНК будет сообщать клеткам устойчивость к ампицилину и не даст устойчивости к эритромицину. Второй фрагмент из-за небольшой длины не может замкнуться в кольцо. Второй вариант лигирования приводит к сшиванию липких концов двух фрагментов. Он происходит примерно в 10 раз реже, а после сшивки вторая пара липких концов скорее всего также, как и исходный фрагмент замкнётся в кольцо. Таких колец из пары фрагментов может образоваться 4 вида: димеры большого фрагмента в двух разных ориентациях (правый конец с левым концом второго фрагмента и левый конец с правым концом второго фрагмента или правый с правым и левый с левым) и соединения большого и малого фрагмента в двух разных ориентациях (вариант исходной плазмиды и инверсия малого фрагмента). Из них только в варианте исходной плазмиды восстанавливается устойчивость к эритромицину. Линейная молекула, образованная сшиванием двух фрагментов, может присоединить ещё один фрагмент с ещё в 10 раз меньшей частотой. Такие фрагменты в дальнейшем будут циклизоваться в плазмиды трёх размеров: из трёх больших фрагментов, из двух больших и одного малого и одного большого и двух малых. Три малых фрагмента дадут короткую последовательность, которая не сможет замкнуться в кольцо и существовать в клетке. В каждом размерном классе будет несколько вариантов с разной ориентацией фрагментов. Только в одном из них восстановится ген устойчивости к эритромицину: правый конец большого фрагмента соединяется с левым концом малого фрагмента, а правый конец малого фрагмента – с левым концом второго большого фрагмента, а оставшиеся концы двух больших фрагментов соединяются с образованием кольцевой плазмиды длиной 8363 пары нуклеотидов. Доля таких молекул будет менее 1% всех плазмид. Вероятность образования плазмид из 4 и более фрагментов ещё на порядок ниже и их обнаружение при данном числе полученных трансформированных клеток нереально. А. Так как расщепление рестриктазой не затрагивает ген устойчивости к ампицилину, все клетки, в результате трансформации получившие любую плазмиду, будут устойчивы к ампицилину и вырастут на среде с этим антибиотиком. Таким образом из 33506 выросших колоний плазмиду получили 578, выросших на ампицилине. Эффективность трансформации представляет долю трансформированных клеток от общего их числа, т.е. 573 : 51366 × 100% = 1.12% Б. На эритромицине могут вырасти только те клетки, в которые попали плазмиды, в которых в результате лигирования восстановится последовательность нуклеотидов в гене устойчивости к этому антибиотику, расщеплённому рестриктазой. Остальные плазмиды, полученные по приведённой методике, будут содержать либо ген с выщепленным коротким фрагментом, что приведёт либо к утрате стартового кодона (если обозначенный зелёным цветом кодон является стартовым), либо к сдвигу рамки считывания (т.к. число удалённых нуклеотидов не кратно трём), либо, при инверсии короткого фрагмента, к появлению стоп-кодонов т.е. прекращению синтеза белка. Таким образом большинство полученных плазмид не обеспечат устойчивости к эритромицину. В. Рост на эритромицине могут обеспечить только плазмиды, несущие восстановленную последовательность гена устойчивости. Такие плазмиды могли образоваться из одного большого и одного малого фрагмента (4200 пар, исходная плазмида)) или из двух больших и одного малого (8363 пары, начало и конец гена из разных копий большого фрагмента). Г. Получается 1 размер из одного большого фрагмента, два размерных класса из двух фрагментов и три размерных класса из трёх фрагментов, то есть 6 размерных классов. (В реальности различить по длине плазмиды, отличающиеся на длину малого фрагмента, т.е. менее чем на 0,5%, невозможно. Поэтому в эксперименте, например на электрофореграмме, будут видны лишь три размерных класса, соответствующие 1, 2 или 3 копиям большого фрагмента.)
При сшивании липких концов ДНК-лигазой наиболее часто будут соединяться концы этой молекулы и образовываться кольцо длиной 4163 нуклеотида. Такая ДНК будет сообщать клеткам устойчивость к ампицилину и не даст устойчивости к эритромицину. Второй фрагмент из-за небольшой длины не может замкнуться в кольцо. Второй вариант лигирования приводит к сшиванию липких концов двух фрагментов. Он происходит примерно в 10 раз реже, а после сшивки вторая пара липких концов скорее всего также, как и исходный фрагмент замкнётся в кольцо. Таких колец из пары фрагментов может образоваться 4 вида: димеры большого фрагмента в двух разных ориентациях (правый конец с левым концом второго фрагмента и левый конец с правым концом второго фрагмента или правый с правым и левый с левым) и соединения большого и малого фрагмента в двух разных ориентациях (вариант исходной плазмиды и инверсия малого фрагмента). Из них только в варианте исходной плазмиды восстанавливается устойчивость к эритромицину. Линейная молекула, образованная сшиванием двух фрагментов, может присоединить ещё один фрагмент с ещё в 10 раз меньшей частотой. Такие фрагменты в дальнейшем будут циклизоваться в плазмиды трёх размеров: из трёх больших фрагментов, из двух больших и одного малого и одного большого и двух малых. Три малых фрагмента дадут короткую последовательность, которая не сможет замкнуться в кольцо и существовать в клетке. В каждом размерном классе будет несколько вариантов с разной ориентацией фрагментов. Только в одном из них восстановится ген устойчивости к эритромицину: правый конец большого фрагмента соединяется с левым концом малого фрагмента, а правый конец малого фрагмента – с левым концом второго большого фрагмента, а оставшиеся концы двух больших фрагментов соединяются с образованием кольцевой плазмиды длиной 8363 пары нуклеотидов. Доля таких молекул будет менее 1% всех плазмид. Вероятность образования плазмид из 4 и более фрагментов ещё на порядок ниже и их обнаружение при данном числе полученных трансформированных клеток нереально. А. Так как расщепление рестриктазой не затрагивает ген устойчивости к ампицилину, все клетки, в результате трансформации получившие любую плазмиду, будут устойчивы к ампицилину и вырастут на среде с этим антибиотиком. Таким образом из 33506 выросших колоний плазмиду получили 578, выросших на ампицилине. Эффективность трансформации представляет долю трансформированных клеток от общего их числа, т.е. 573 : 51366 × 100% = 1.12% Б. На эритромицине могут вырасти только те клетки, в которые попали плазмиды, в которых в результате лигирования восстановится последовательность нуклеотидов в гене устойчивости к этому антибиотику, расщеплённому рестриктазой. Остальные плазмиды, полученные по приведённой методике, будут содержать либо ген с выщепленным коротким фрагментом, что приведёт либо к утрате стартового кодона (если обозначенный зелёным цветом кодон является стартовым), либо к сдвигу рамки считывания (т.к. число удалённых нуклеотидов не кратно трём), либо, при инверсии короткого фрагмента, к появлению стоп-кодонов т.е. прекращению синтеза белка. Таким образом большинство полученных плазмид не обеспечат устойчивости к эритромицину. В. Рост на эритромицине могут обеспечить только плазмиды, несущие восстановленную последовательность гена устойчивости. Такие плазмиды могли образоваться из одного большого и одного малого фрагмента (4200 пар, исходная плазмида)) или из двух больших и одного малого (8363 пары, начало и конец гена из разных копий большого фрагмента). Г. Получается 1 размер из одного большого фрагмента, два размерных класса из двух фрагментов и три размерных класса из трёх фрагментов, то есть 6 размерных классов. (В реальности различить по длине плазмиды, отличающиеся на длину малого фрагмента, т.е. менее чем на 0,5%, невозможно. Поэтому в эксперименте, например на электрофореграмме, будут видны лишь три размерных класса, соответствующие 1, 2 или 3 копиям большого фрагмента.)
pазбирался: Надежда | обсудить разбор | оценить
По принципу комплементарности строим