Стоп кодон – полное руководство
Определение
Стоп-кодон является генетический код что сигнализирует об окончании производства белка внутри клетка, как точка в конце предложения. Три стоп-кодона нуклеотид базовые триплеты, которые играют важную роль во внутриклеточном синтезе белка; физиологические и / или анатомические изменения возможны, если стоп-кодон находится в неправильном положении на цепи ДНК или РНК или если кодовая последовательность изменена.

Без стоп-кодонов организм не может производить конкретные белки. Новая полипептидная (белковая) цепь будет просто расти и расти до тех пор, пока клетка не лопнет или не исчезнет. аминокислоты добавить к этому. И стартовые, и стоп-кодоны в ДНК и РНК, как и предполагают их названия, предоставляют инструкции по запуску и остановке, которые регулируют длину полипептидной цепи. Каждая цепь является результатом отдельных аминокислот, связанных в определенном порядке, как показано ниже.

Все кодоны состоят из трех нуклеотидных оснований и названы в соответствии с порядком этих оснований – например, стоп-кодон TAG говорит нам, что он состоит из тимин затем аденин, затем гуанин. Чтобы по-настоящему понять важность стоп-кодона, полезно освежить наши знания о конструкции ДНК и синтезе белка.


Цитогенное местоположение говорит ученым, где найти различные инструкции по производству белка. Также важно помнить, что, хотя каждый клеточное ядро содержит инструкции для выработки полнофункционального организма, большинство генов экспрессируются (активируются) только в определенных тканях; KRT-9 экспрессируется в клетках кожи ладоней и подошв, а печень клеточное ядро также содержит инструкции по производству кератина 9, но ген не экспрессируется.

Хотя многие источники говорят о зеркальной копии мРНК, они не всегда упоминают, что это зеркальная копия зеркальной копии и, следовательно, точная копия кодирующей цепи ДНК. Это легче понять, если учесть, что ДНК состоит из двух отдельных цепей – кодирующей (смысловой) цепочки, которая проходит в одну сторону, и шаблонной (антисмысловой) цепочки, которая идет антипараллельно ей. Если, например, смысловая цепь проходит слева направо, антисмысловая цепь проходит справа налево. Если смысловая цепь содержит последовательность AAAGCC, антисмысловая цепь будет состоять из нуклеотидов-партнеров, идущих в противоположном направлении: GGCTTT. РНК затем транскрибирует (копирует) код антисмысловой цепи антипараллельно, то есть слева направо – точно так же, как смысловая цепь. Это означает, что код РНК будет AAAGCC – точно такой же, как код антисмысловой цепи ДНК. Существует только одна разность потенциалов – партнером аденина в ДНК является тимин, а в РНК тимин заменяется урацилом.
Как только эти присоединения были сделаны во время процесса транскрипции ДНК, цепь РНК переименовывается в мессенджер РНК или мРНК.
![]()
Ученые согласны с тем, что в генетическом коде человека есть три стоп-кодона – также называемые нонсенс-кодонами или терминирующими кодонами. Это TAG, TAA и TGA (ДНК) и UAG, UAA и UGA (РНК). Опять же, TAG, TAA и TGA не действуют как стоп-кодоны во время транскрипции, но копируются (заменяя тимин на урацил) РНК. Стоп-кодоны не кодируют аминокислоту и не относятся к некодирующей группе генов, но являются отдельным объектом. Их распознавание намного проще, чем распознавание стартового кодона. В то время как стартовый кодон также кодирует аминокислоту под названием метионин, аминокислоты стоп-кодона не существуют; их триплетные нуклеотидные последовательности не кодируют часть полипептидной цепи, а действуют только для завершения процессов транскрипции и трансляции.

После того как стартовый кодон мРНК обнаружен, наступает время для переноса РНК доставлять нужные аминокислоты в том же порядке, что и связанные с ними нуклеотидные триплеты. каждый тРНК несет аминокислоту, которая соответствует кодону на мРНК. Трансферная РНК или тРНК «читает» кодоны мРНК, поэтому этот этап синтеза белка называется трансляцией. Именно на этапе трансляции запускаются и останавливаются функции кодонов.
![]()


Какие три стоп-кодона?
Три стоп-кодона – это TAG, TAA и TGA в смысле ДНК и UAG, UAA и UGA в мРНК.
TAG и UAG называются янтарными стоп-кодонами; TAA и UAA известны как стоп-кодоны охры, а TGA и UGA – названия опаловых стоп-кодонов (или янтарных стоп-кодонов). Код янтарного цвета приписывается имени ученого, который первым его обнаружил; другие цвета просто продолжают эту цветовую тему. Стоп-кодоны также называют нонсенс-кодонами или терминирующими кодонами, первый из этих терминов, потому что стоп-кодоны никогда не кодируют аминокислоты, а второй – из-за функции стоп-кодонов.
Стоп кодонов мутаций
Мутации стоп-кодонов могут легко возникнуть, особенно если учесть длину генома и тысячи различных нуклеотидных триплетов. Как процессы транскрипции, так и трансляции подвержены широкому кругу потенциальных ошибок, которые могут или не могут привести к анатомическим и физиологическим изменениям. вставка неправильного нуклеотида в ген KRT-9 у членов семьи Было обнаружено, что уже предрасположенные к этому заболеванию способствуют развитию кожного заболевания, известного как эпидермолитическая пальмоплантарная кератодерма.

В то время как все виды мутаций происходят во время транскрипции ДНК в мРНК, мРНК копирует только то, что написано, даже не понимая этого. В течение периода, когда мРНК не контактирует с рибосомой, даже множественные мутации не будут вызывать эффекта. Эффекты видны только тогда, когда измененный код транслируется в дефектный белок. Именно поэтому большинство мутаций помечены как часть процесса трансляции, когда отредактированный код может производить или не производить другую аминокислоту. Тот факт, что большинство аминокислот соответствуют шести различным нуклеотидным триплетам, означает, что существует вероятность того, что даже при наличии мутации будет продуцироваться один и тот же белок. Мы обычно связываем генетические мутации с болезнью; однако они также несут ответственность за успешную эволюцию. Генетические мутации помогают организмам адаптироваться к окружающей среде.
Существуют различные формы генетической мутации. Мутации удаления не копируют определенные части генома и, таким образом, изменяют порядок нуклеотидов. Одна база или несколько баз могут быть полностью пропущены. Мутации вставки добавляют один или несколько нуклеотидов, а также изменяют порядок генетического кода. Заместительные мутации (молчащие, миссенс и нонсенс) заменяют один нуклеотид (не несколько нуклеотидов) другим основанием, и это может заменить или не заменить другую аминокислоту в полипептидной цепи. Если тот же белок продуцируется, даже в присутствии мутации, он называется тихая мутация, В некоторых случаях целый участок ДНК может меняться между двумя нитями – это называется транслокацией.
Если в полипептидную цепь добавлена другая аминокислота, которая может изменить или не изменить ее функцию, причиной является миссенс мутация, Если замещение создает стоп-кодон путем изменения кода нуклеотидного триплета, который соответствует аминокислоте, это называется нонсенс-мутацией. На рисунке ниже показаны три типа мутаций: A – нонсенс-мутация, B – инсерционная мутация, а C и D – делеционные мутации.
Стоп-кодон
Стоп-кодоны выполняют важную функцию завершения (терминацию) сборки полипептидной цепи и также называются терминаторными кодонами. Некоторые из них вызывают обязательное прекращение синтеза, другие являются условными.
Кроме того, стоп-кодон, как кодон, при котором не происходит включения аминокислоты в белок, ещё называют бессмысленным кодоном или нонсенс-кодоном.
Так, кодон UAG (Янтарь) — условный терминаторный кодон и супрессируемые Amber-мутации вызывают преждевременную терминацию трансляции (условно летальные мутации).
Сквозная трансляция может проходить через кодоны UAG (Янтарь) и UGA (Опал), но не через кодон UAA (Охра).
Кодоны UAA и UAG в митохондриальной ДНК вызывают безусловное прекращение трансляции.
Некоторые мРНК в действительности содержат два тандемных терминаторных кодона — часто это кодоны различного типа на конце кодирующей последовательности.
Связанные понятия
70 нуклеотидов, иногда 80-100 нуклеотидов), богатый цитозином и бедными в гуанином, называемый ро-утилизационным сайтом (rut), синтезируемым в РНК, выше фактической последовательности терминатора. Были выявлены несколько связывающих последовательностей.
ДНК-метилтрансфера́зы (ДНК-метилазы, англ. DNA methyltransferase, DNA MTase, DNMT) — группа ферментов, катализирующих метилирование нуклеотидных остатков в составе ДНК. Активность метилтрансфераз, заключающаяся в переносе метильных (CH3—) групп на азотистое основание цитозин в составе ДНК, ведет к изменению свойств ДНК, при этом изменяется активность, функции соответствующих генов, а также пространственная структура нуклеиновой кислоты (конформация).
Неоднозначность стоп-кодонов

Языки людей неоднозначны, многие слова в них имеют не одно, а несколько значений. Обычно мы улавливаем смысл исходя из контекста сказанного. У клеток свой язык, который они используют при синтезе белка. Это генетический код, состоящий из 64 нуклеотидных триплетов (кодонов), 61 из которых, смысловые, кодируют определенные аминокислоты, а три стоп-кодона вызывают завершение синтеза белковой молекулы (трансляции). Но кодоны, оказывается, могут иметь разный смысл, и, чтобы его понять, приходится, опять-таки, ориентироваться на контекст.
Про инвариантность генетического кода написано в учебниках, однако специалисты знают, что это не так. Впервые его неоднозначность обнаружили еще в 1985 году у инфузорий, а затем у зеленых водорослей и дипломонад, а также в митохондриальных геномах. В этих случаях стоп-кодоны не только терминируют синтез белка, но и кодируют в нем какую-нибудь аминокислоту. И как, спрашивается, рибосомы различают, в каких случаях этот кодон следует прочесть, а в каких — на нем остановиться? Ответ на этот вопрос искали специалисты Бернского и Пизанского университетов под руководством бернского профессора Мариуша Новацки (Mariusz Nowacki).
Исследователи проанализировали обширную базу данных транскриптомов (совокупностей всех молекул РНК) морских одноклеточных эукариот (Marine Microbial Eukaryote Transcriptome Sequencing Project, MMETSP), чтобы обнаружить и классифицировать переназначенные кодоны. Три переназначенных стоп-кодона они нашли в геномах нескольких видов инфузорий: UAA и UAG кодируют глютамин, цистеин или тирозин, а UGA — триптофан. Каждому смысловому кодону соответствует тРНК, подносящая определенную аминокислоту, а со стоп-кодонами взаимодействуют особые белки — факторы терминации трансляции. Таким образом, за каждый из переназначенных кодонов конкурируют по крайней мере одна тРНК и фактор терминации трансляции eRF1.
У большинства исследованных видов инфузорий с переназначенными стоп-кодонами оставался хотя бы один инвариантный, но у Condylostoma magnum и Parduzcia sp. переназначенными оказались все три: UAA и UAG кодируют глютамин, UGA — триптофан. Именно с этими двумя видами исследователи и продолжили работу. Изучение начали с последовательности гистона Н4 — одной из самых консервативных. Она обычно заканчивается стоп-кодонами UAG или UGA. Эксперименты показали, что рибосома проскакивает эту позицию крайне редко, с частотой менее 1,8%, обычно в этом месте происходит терминация. Однако в тех случаях, когда стоп-кодоны находятся в середине кодирующей последовательности, рибосома воспринимает их как смысловые. Ученые нашли тРНК, которая распознает UAA и UAG. Досрочной терминации трансляции при этом не происходит.
Как можно различать смысл неоднозначных кодонов? Исследователи предложили две гипотезы: либо рядом с кодонами находятся специфические последовательности, позволяющие тРНК или eRF1 сделать правильный выбор, или же значение кодона определяется его положением в молекуле РНК. Если стоп-кодон находится в конце молекулы, регуляторные белки воспринимают его как сигнал остановки. Подходящих последовательностей-маркеров, позволяющих определить значение кодона, ученые не обнаружили, поэтому сосредоточились на второй гипотезе.
Молекулы мРНК, с которых в рибосоме считываются белковые молекулы, имеют характерное строение. Их кодирующая последовательность оканчивается стоп-кодоном, за ним следует короткая 3’-некодирующая область и несколько адениловых остатков — поли(А)-хвост (см. рисунок). 3’-некодирующая область может быть настолько короткой, что поли(А)-хвост порой находится практически вплотную к стоп-кодону, поэтому ученые полагают, что именно положение стоп-кодона относительно поли(А)-хвоста позволяет определить его значение. Если хвост совсем рядом, надо ставить «точку»; стоп-кодоны, расположенные чуть дальше от поли(А), в 24–66 нуклеотидах от последнего кодона, читаются как смысловые.

Если какой-либо смысловой кодон в определенном положении часто читают как «стоп», отбор его оттуда уберет, потому что ошибка обойдется слишком дорого. Если гипотеза о том, что значение стоп-кодона зависит от его положения относительно 3’-конца молекулы, верна, подобные ошибки должны чаще всего происходить в тех случаях, когда смысловые стоп-кодоны расположены не в самом конце молекулы, но поблизости от него, и приводить к досрочной терминации. В таком случае в ходе эволюции они должны были исчезнуть. И действительно, в конце кодирующей области мРНК, перед настоящим стоп-кодоном, его смысловых собратьев нет. Ни для каких других кодонов таких позиционных эффектов не отмечено.
Итак, Мариуш Новацки и его коллеги предложили модель, согласно которой стоп-кодоны в молекулах мРНК инфузорий C. magnum и Parduczia sp. по умолчанию читаются как смысловые, а не служат стоп-сигналом. Терминация трансляции происходит, когда стоп-кодон расположен в самом конце молекулы. Исследователи пришли к выводу, что на срабатывание кодона как стоп-сигнала влияет близость поли(А)-хвоста и белков, с ним взаимодействующих. Особую роль они отводят белку РАВР.
Поскольку стоп-кодоны распределены по всей длине гена и благополучно транслируются, речь идет не о мутациях, а об изменении генетического кода. Бросается в глаза, что такие изменения произошли только у инфузорий, у других 265 эукариотических видов из MMETSP их нет.
По мнению исследователей, неоднозначность генетического кода инфузорий отражает первоначальную неоднозначность кода, когда каждый кодон имел несколько значений. Возникновение в ходе эволюции очень короткой 3’-нетранслируемой области и поли(А)-хвоста позволило переобозначить стоп-кодоны, использовать их в качестве сигнала терминации. Такое контекстное прочтение делает генетический код устойчивее к мутациям, превращающим значащие кодоны в стоп-кодоны. Если такая мутация произойдет в геноме с инвариантным кодом, она приведет к образованию короткого мутантного белка и преждевременной терминации трансляции. Но когда подобное случится в неоднозначном геноме инфузории, стоп-кодон будет прочитан, поскольку находится на месте смыслового, а для терминации трансляции нужен не только определенный триплет, но и его особое положение.
По мнению профессора Новацки и его соавторов, предки инфузорий долгое время прекрасно существовали с неоднозначными генетическими кодами. Нынешний инвариантный генетический код вовсе не последнее слово эволюции: генетические коды иногда изменяются.
Генетический код клетки
Ранее мы подчёркивали, что нуклеотиды имеют важную для формирования жизни на Земле особенность ― при наличии в растворе одной полинуклеотидной цепочки спонтанно происходит процесс образования второй (параллельной) цепочки на основании комплементарного соединения родственных нуклеотидов. Одинаковое число нуклеотидов, в обоих цепочках и их химическое родство, является непременным условием для осуществления такого рода реакций.
Однако при синтезе белка, когда информация с иРНК реализуется в структуру белка никакой речи о соблюдении принципа комплементарности идти не может. Это связано с тем, что в иРНК, и в синтезированном белке различно не только число мономеров, но и, что особенно важно, отсутствует структурное сходство между ними (с одной стороны нуклеотиды, с другой аминокислоты). Понятно, что в этом случае возникает необходимость создания нового принципа точного перевода информации с полинуклеотида в структуру полипептида. В эволюции такой принцип был создан и в его основу был заложен генетический код.
Генетический код ― это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны, соответствующие аминокислотам в белке.
Свойства генетического кода
Генетический код имеет несколько свойств:
Следует отметить, что некоторые авторы предлагают ещё и другие свойства кода, связанные с химическими особенностями входящих в код нуклеотидов или с частотой встречаемости отдельных аминокислот в белках организма и т.д. Однако эти свойство вытекают из вышеперечисленных, поэтому там мы их и рассмотрим.
Триплетность
Генетический код, как и многое сложно организованные система имеет наименьшую структурную и наименьшую функциональную единицу. Триплет ― наименьшая структурная единица генетического кода. Состоит она из трёх нуклеотидов. Кодон ― наименьшая функциональная единица генетического кода. Как правило, кодонами называют триплеты иРНК. В генетическом коде кодон выполняет несколько функций. Во-первых, главная его функция заключается в том, что он кодирует одну аминокислоту. Во-вторых, кодон может не кодировать аминокислоту, но, в этом случае, он выполняет другую функцию (см. далее). Как видно из определения, триплет ― это понятие, которое характеризует элементарную структурную единицу генетического кода (три нуклеотидов). Кодон ― характеризует элементарную смысловую единицу генома ― три нуклеотида определяют присоединение к полипептидной цепочки одной аминокислоты.
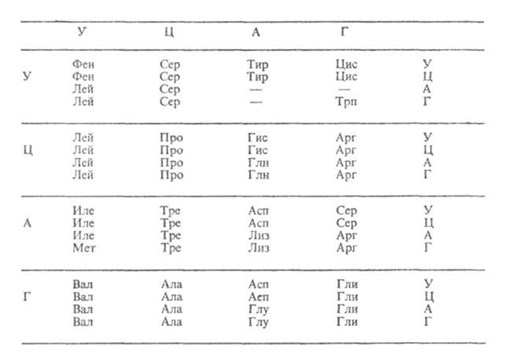
Элементарную структурную единицу вначале расшифровали теоретически, а затем её существование подтвердили экспериментально. И действительно, 20 аминокислот невозможно закодировать одним или двумя нуклеотидом т.к. последних всего 4. Три нуклеотида из четырёх дают 43 = 64 варианта, что с избытком перекрывает число имеющихся у живых организмах аминокислот (см.табл. 1).
Представленные в таблице 64 сочетания нуклеотидов имеют две особенности. Во-первых, из 64 вариантов триплетов только 61 являются кодонами и кодируют какую-либо аминокислоту, их называют смысловые кодоны. Три триплета не кодируют.
Таблица 1.
Как пользоваться этой таблицей, смотрите в этом видео:
Стоп-кодоны
Кодоны информационной РНК и соответствующие им аминокислоты являются стоп-сигналами, обозначающие конец трансляции. Таких триплетов три ― УАА, УАГ, УГА, их ещё называют «бессмысленные» (нонсенс кодоны). В результате мутации, которая связана с заменой в триплете одного нуклеотида на другой, из смыслового кодона может возникнуть бессмысленный кодон. Такой тип мутации называют нонсенс-мутация. Если такой стоп-сигнал сформировался внутри гена (в его информационной части), то при синтезе белка в этом месте процесс будет постоянно прерываться ― синтезироваться будет только первая (до стоп-сигнала) часть белка. У человека с такой патологией будет ощущаться нехватка белка и возникнут симптомы, связанные с этой нехваткой. Например, такого рода мутация выявлена в гене, кодирующем бета-цепь гемоглобина. Синтезируется укороченная неактивная цепь гемоглобина, которая быстро разрушается. В результате формируется молекула гемоглобина, лишённая бета-цепи. Понятно, что такая молекула вряд ли будет полноценно выполнять свои обязанности. Возникает тяжёлое заболевания, развивающееся по типу гемолитической анемии (бета-ноль талассемия, от греческого слова «Таласа» ― Средиземное море, где эта болезнь впервые обнаружена).
Механизм действия стоп-кодонов отличается от механизма действия смысловых кодонов. Это следует из того, что для всех кодоны, кодирующие аминокислоты, найдены соответствующие тРНК. Для нонсенс-кодонов тРНК не найдены. Следовательно, в процессе остановки синтеза белка тРНК не принимает участие.
Кодон АУГ (у бактерий иногда ГУГ) не только кодируют аминокислоту метионин и валин, но и является инициатором трансляции.
Неканонические значения кодонов
По крайней мере у 16 типов организмов генетический код отличается от канонического. Например многие виды зелёных водорослей Acetabularia транслируют стандартные стоп-кодоны UAG и UAA в аминокислоту глицин, а гриб Candida интерпретирует РНК-кодон CUG не как лейцин, а как серин. А у митохондрий пекарских дрожжей (Saccharomyces cerevisiae) четыре из шести кодонов, обычно транслирующихся в лейцин, кодируют треонин.
Существование таких вариаций свидетельствует о возможной эволюции генетического кода.
Представители всех трёх доменов живых организмов иногда прочитывают стандартный стоп-кодон UGA как 21-ю аминокислоту селеноцистеин, не относящуюся к 20 стандартным. Селеноцистеин образуется при химической модификации серина на стадии, когда последний ещё не отсоединился от тРНК в составе рибосомы.
Аналогично у представителей двух доменов (архебактерий и бактерий) стоп-кодон UAG прочитывается как 22-я аминокислота пирролизин.
Вырожденность, или избыточность
61 из 64 триплетов кодируют 20 аминокислот. Такое трёхразовое превышение числа триплетов над количеством аминокислот позволяет предположить, что в переносе информации могут быть использованы два варианта кодирования. Во-первых, не все 64 кодона могут быть задействованы в кодировании 20 аминокислот, а только 20 и, во-вторых, аминокислоты могут кодироваться несколькими кодонами. Исследования показали, что природа использовала последний вариант.
Код, при котором одна аминокислота кодируется несколькими триплетами, называется вырожденным или избыточным. Почти каждой аминокислоте соответствует несколько кодонов. Так, аминокислота лейцин может кодироваться шестью триплетами — УУА, УУГ, ЦУУ, ЦУЦ, ЦУА, ЦУГ. Валин кодируется четырьмя триплетами, фенилаланин — двумя и только триптофан и метионин кодируются одним кодоном. Свойство, которое связано с записью одной и той же информации разными символами носит название вырожденность.
Число кодонов, предназначенных для одной аминокислоты, хорошо коррелируется с частотой встречаемости аминокислоты в белках. И это, скорее всего, не случайно. Чем больше частота встречаемости аминокислоты в белке, тем чаще представлен кодон этой аминокислоты в геноме, тем выше вероятность его повреждения мутагенными факторами. Поэтому понятно, что мутированный кодон имеет больше шансов кодировать туже аминокислоту при высокой его вырожденности. С этих позиций вырожденность генетического кода является механизмом защищающим геном человека от повреждений.
Необходимо отметить, что термин вырожденность используется в молекулярной генетике и в другом смысле. Так основная часть информации в кодоне приходится на первые два нуклеотида, основание в третьем положении кодона оказывается малосущественным. Этот феномен называют «вырожденностью третьего основания». Последняя особенность сводит до минимума эффект мутаций. Например, известно, что основной функцией эритроцитов крови является перенос кислорода от легких к тканям и углекислого газа от тканей к легким. Осуществляет эту функцию дыхательный пигмент — гемоглобин, который заполняет всю цитоплазму эритроцита. Состоит он из белковой части ― глобина, который кодируется соответствующим геном. Кроме белка в молекулу гемоглобина входит ген, содержащий железо. Мутации в глобиновых генах приводят к появлению различных вариантов гемоглобинов. Чаще всего мутации связаны с заменой одного нуклеотида на другой и появлением в гене нового кодона, который может кодировать новую аминокислоту в полипептидной цепи гемоглобина. В триплете, в результате мутации может быть заменён любой нуклеотид ― первый, второй или третий.
Мутации в гемоглобине
Известно несколько сотен мутаций, затрагивающих целостность генов глобина. Около 400 из них связаны с заменой единичных нуклеотидов в гене и соответствующей аминокислотной заменой в полипептиде. Из них только 100 замен приводят к нестабильности гемоглобина и различного рода заболеваниям от легких до очень тяжелых. 300 (примерно 64%) мутаций-замен не влияют на функцию гемоглобина и не приводят к патологии. Одной из причин этого является упомянутая выше «вырожденность третьего основания», когда замена третьего нуклеотида в триплете, кодирующем серин, лейцин, пролин, аргинин и некоторые другие аминокислоты приводит к появлению кодона-синонима, кодирующего ту же аминокислоту. Фенотипически такая мутация не проявится. В отличие от этого любая замена первого или второго нуклеотида в триплете в 100 % случаях приводит к появлению нового варианта гемоглобина. Но и в этом случае тяжёлых фенотипических нарушений может и не быть. Причиной этому является замена аминокислоты в гемоглобине на другую сходную с первой по физико-химическим свойствам. Например, если аминокислота, обладающая гидрофильными свойствами, заменена на другую аминокислоту, но с такими же свойствами.
Гемоглобин состоит из железопорфириновой группы гема (к ней и присоединяются молекулы кислорода и углекислоты) и белка — глобина. Гемоглобин взрослого человека (НвА) содержит две идентичные a-цепи и две b-цепи. Замена в гене, кодирующем b-цепь гемоглобина первого или второго нуклеотида практически всегда приводит к появлению в белка новых аминокислот, нарушению функций гемоглобина и тяжёлым последствия для больного. Например, замена “Ц” в одном из триплетов ЦАУ (гистидин) на “У” — приведет к появлению нового триплета УАУ, кодирующего другую аминокислоту — тирозин. Фенотипически это проявится в тяжёлом заболевании.
Гистидин, тирозин
Аналогичная замена в 63 положении b-цепи полипептида гистидина на тирозин приведет к дестабилизации гемоглобина. Развивается заболевание метгемоглобинемия. Замена, в результате мутации, глутаминовой кислоты на валин в 6-м положении b-цепи является причиной тяжелейшего заболевания — серповидно-клеточной анемии. Не будем продолжать печальный список.
Глутаминовая кислота и лизин
Отметим только, что при замене первых двух нуклеотидов может появится аминокислота по физико-химическим свойствам похожая на прежнюю. Так, замена 2-го нуклеотида в одном из триплетов, кодирующего глутаминовую кислоту (ГАА) в b-цепи на “У” приводит к появлению нового триплета (ГУА), кодирующего валин, а замена первого нуклеотида на “А” формирует триплет ААА, кодирующий аминокислоту лизин. Глутаминовая кислота и лизин сходны по физико-химическим свойствам — они обе гидрофильны. Валин — гидрофобная аминокислота. Поэтому, замена гидрофильной глутаминовой кислоты на гидрофобный валин, значительно меняет свойства гемоглобина, что, в конечном итоге, приводит к развитию серповидноклеточной анемии, замена же гидрофильной глутаминовой кислоты на гидрофильный лизин в меньшей степени меняет функцию гемоглобина — у больных возникает легкая форма малокровия.
Урацил и цитозин
В результате замены третьего основания новый триплет может кодировать туже аминокислоты, что и прежней. Например, если в триплете ЦАУ урацил был заменён на цитозин и возник триплет ЦАЦ, то практически никаких фенотипических изменений у человека выявлено не будет. Это понятно, т.к. оба триплета кодируют одну и туже аминокислоту ― гистидин.
В заключении уместно подчеркнуть, что вырожденность генетического кода и вырожденность третьего основания с общебиологических позиция являются защитными механизмами, которые заложены в эволюции в уникальной структуре ДНК и РНК.
Однозначность
Каждый триплет (кроме бессмысленных) кодирует только одну аминокислоту. Таким образом, в направлении кодон ― аминокислота генетический код однозначен, в направлении аминокислота ― кодон ― неоднозначен (вырожденный).
И в этом случае необходимость однозначности в генетическом коде очевидна. При другом варианте при трансляции одного и того же кодона в белковую цепочку встраивались бы разные аминокислоты и в итоге формировались белков с различной первичной структурой и разной функцией. Метаболизм клетки перешёл бы в режим работы «один ген ― несколько полипептидов». Понятно, что в такой ситуации регулирующая функция генов была бы полностью утрачена.
Полярность
Считывание информации с ДНК и с иРНК происходит только в одном направлении. Полярность имеет важное значение для определения структур высшего порядка (вторичной, третичной и т.д.). Ранее мы говорили о том, что структуры низшего порядка определяют структуры более высшего порядка. Третичная структура и структуры более высокого порядка у белков, формируются сразу же как только синтезированная цепочка РНК отходит от молекулы ДНК или цепочка полипептида отходит от рибосомы. В то время, когда свободный конец РНК или полипептида приобретает третичную структуру, другой конец цепочки ещё продолжает синтезироваться на ДНК (если транскрибируется РНК) или рибосоме (если транскрибируется полипептид).
Поэтому однонаправленный процесс считывания информации (при синтезе РНК и белка) имеет существенное значение не только для определения последовательности нуклеотидов или аминокислот в синтезируемом веществе, но для жёсткой детерминации вторичной, третичной и т.д. структур.
Неперекрываемость
Код может быть перекрывающимся и не перекрывающимся. У большинства организмов код не перекрывающийся. Перекрывающийся код найден у некоторых фагов.
Сущность не перекрывающего кода заключается в том, что нуклеотид одного кодона не может быть одновременно нуклеотидом другого кодона. Если бы код был перекрывающим, то последовательность из семи нуклеотидов (ГЦУГЦУГ) могла кодировать не две аминокислоты (аланин-аланин) как в случае с не перекрывающимся кодом, а три (если общим является один нуклеотид) или пять (если общими являются два нуклеотида). В последних двух случаях мутация любого нуклеотида привела бы к нарушению в последовательности двух, трёх и т.д. аминокислот.
Однако установлено, что мутация одного нуклеотида всегда нарушает включение в полипептид одной аминокислоты. Это существенный довод в пользу того, что код является не перекрывающимся. Неперекрываемость генетического кода связана с ещё одним свойством ― считывание информации начинается с определённой точки ― сигнала инициации. Таким сигналом инициации в иРНК является кодон, кодирующий метионин АУГ. Следует отметить, что у человека всё-таки имеется небольшое число генов, которые отступают от общего правила и перекрываются.
Компактность
Между кодонами нет знаков препинания. Иными словами триплеты не отделены друг от друга, например, одним ничего не значащим нуклеотидом. Отсутствие в генетической коде «знаков препинания» было доказано в экспериментах.
Универсальность
Код един для всех организмов, живущих на Земле. Прямое доказательство универсальности генетического кода было получено при сравнении последовательностей ДНК с соответствующими белковыми последовательностями. Оказалось, что во всех бактериальных и эукариотических геномах используется одни и те же наборы кодовых значений. Есть и исключения, но их не много.
Первые исключения из универсальности генетического кода были обнаружены в митохондриях некоторых видов животных. Это касалось кодона терминатора УГА, который читался так же как кодон УГГ, кодирующий аминокислоту триптофан. Были найдены и другие более редкие отклонения от универсальности.
Краткие сведения
Генетический код ― это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны, соответствующие аминокислотам в белке. Генетический код имеет несколько свойств.